Alan Koh-Siong Nga1,
Lai-Yeng Tho2,
Cheng-Hoe Lim2,
Chan-Kiang Lim2,
Yee-How Say1 ![]()
For correspondence:- Yee-How Say Email: sayyh@utar.edu.my Tel:+6054688888
Received: 26 August 2016 Accepted: 19 November 2016 Published: 21 December 2016
Citation: Nga AK, Tho L, Lim C, Lim C, Say Y. Evaluation of neuroprotective properties of two synthetic prenylated xanthone analogues against paraquat and 6-hydroxydopamine toxicity in human neuroblastoma SH-SY5Y cells. Trop J Pharm Res 2016; 15(12):2611-2618 doi: 10.4314/tjpr.v15i12.11
© 2016 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To investigate whether two synthetic prenylated xanthone analogues - 1,3,6,8-tetrahydroxy-9H-xanthen-9-one (SX1) and 1,3,6-trihydroxy-2-(3-methylbut-2-enyl)-9H-xanthen-9-one (SX2) - are potential candidates for neuroprotection against paraquat- and 6-hydroxydopamine (OHDA)-induced human neuroblastoma SH-SY5Y cell death.
Methods: SH-SY5Y cells were treated with SX1 and SX2, and the maximum non-toxic dose (MNTD) were obtained by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. MTT assay was also used to assess the ability of MNTD and half MNTD (HMNTD) doses of SX1 and SX2 to protect against the neurotoxicity of 200 µM paraquat and 100 µM 6-OHDA. Intracellular ROS production by SH-SY5Y cells treated or untreated with SX1 or SX2 was measured by dichloro-dihydro-fluorescein diacetate (DCFH-DA) assay.
Results: SX1 and SX2 MNTDs at concentrations of 1850 nM and 105 nM, respectively, did not significantly (p > 0.05) provide neuroprotection against paraquat-induced SH-SY5Y cell death. Only SX2 MNTD and HMNTD significantly (p < 0.05) protected SH-SH5Y cells against 6-OHDA-induced cell death by 10 and 17 % improved cell viability. Although intracellular ROS production was significantly attenuated by SX1 HMNTD and MNTD, this did not improve cell viability against paraquat-induced cell death.
Conclusion: These results reveal that SX2 confers neuroprotection on 6-OHDA-induced SH-SY5Y neurotoxicity. Further investigations to elucidate the detailed molecular mechanisms of neuroprotection by SX2 are warranted.
Introduction
Paraquat and 6-hydroxydopamine (6-OHDA) have been used in the laboratory as prototypical neurotoxins to induce neurodegeneration in both in vivo and in vitro settings. 6-OHDA induces neurotoxicity by producing hydrogen peroxide, superoxide radical, oxidized glutathione and hydroxyl radical during its autoxidation [1]. These oxidative stresses may ultimately result in neurodegeneration by inducing inflammation of neurons and death of neurons in the brain [2]. Paraquat has been considered as a possible etiologic factor in Parkinson's disease. Divalent paraquat ion (PQ2+) in brain is reduced to monovalent paraquat ion (PQ+) by extracellular enzymes such as NADPH-oxidase in microglia, which then serves as a dopamine transporter substrate [3]. Subsequently, PQ+ accumulates in dopaminergic neurons and reestablishes a new redox reaction intracellularly, leading to the generation of intracellular free radicals such as superoxide and dopamine reactive substances. This will eventually lead to dopaminergic neuron cell death as seen in Parkinson’s disease [3].
Several phytochemicals including fatty acids, phenols, flavonoids, saponins, xanthones and terpenes found in natural products are potential neuroprotective compounds due to their anti-oxidation and anti-lipid peroxidation activities [4]. Natural or synthetic xanthones, in view of their unique structural chromophore, have been reported to exhibit a wide range of biological properties including cytotoxicity, anti-inflammatory, antioxidant, antimalarial, antibacterial and antifungal activities [5]. Our previous studies showed that certain synthetic tri- and tetra-oxygenated prenylated xanthones had cytotoxic activities against cervical and breast cancer cell lines [6,7]. A recent finding showed that six newly isolated prenylated xanthones from ethyl acetate soluble extract of Cudrania tricusidata root bark together with six previously identified prenylated xanthones had neuroprotective effects against 6-OHDA-induced cell death in human neuroblastoma SH-SY5Y cells [8]. This provides an early finding that prenylated xanthones could have neuroprotective property against certain drugs or chemicals that induce neurodegeneration like paraquat and 6-OHDA.
Therefore, the objective of this study was to evaluate the neuroprotective property of two selected synthetic prenylated xanthone analogues from our previous studies [6,7], against the toxicities of paraquat and 6-hydroxydopamine in a dopaminergic cell line - human neuroblastoma SH-SY5Y cells.
Methods
Prenylated xanthones
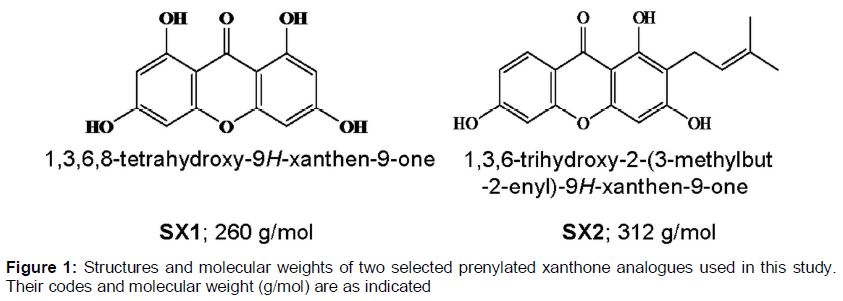
Two selected prenylated xanthone analogues were from our previous studies 1,3,6-trihydroxy-9H-xanthen-9-one (SX1) is compound 1 in [6] and 1,3,6-trihydroxy-2-(3-methylbut-2-enyl)-9H-xanthen-9-one (SX2) is compound 8 in [7]. The structures are shown in and the synthesis processes were as outlined in our previous studies [6,7]. These compounds were selected as they have IC50 of > 60 μM in human breast cancer MDA-MB-231 cell line [6,7]. A higher IC50 of a compound would enable a wider range of dosages to be tested on SH-SY5Y during the determination of Maximum Non-toxic Dose (MNTD).
Cell culture
Human neuroblastoma SH-SY5Y (ATCC® CRL-2266™) was obtained from the American Type Culture Collection (Manassas, VA, U.S.A.). The cells were maintained in Dulbecco’s Modified Eagle’s Medium (DMEM; Thermo Fischer Scientific PA, USA) containing 10 % Fetal Bovine Serum (FBS; Sigma-Aldrich, MO, USA) and 1 % penicillin-streptomycin (Nacalai Tesque, Japan) by incubating at 37 °C in a 5 %(v/v) CO2 humidified incubator.
Determination of maximum non-toxic dose (MNTD) by MTT assay and treatment paradigm
SH-SY5Y human neuroblastoma cells were seeded in 96-well plates at a cell density of 8,000 cells/well in 100 µL of complete growth. Upon attachment the next day, the cells were replaced with serum-free medium (basic medium) on next day for another 24 h to synchronize the growth stage of cells. On the third day, the cells were treated with synthetic prenylated xanthones SX1 and SX2 dissolved in dimethyl sulfoxide (DMSO), at a range of concentrations from 0 to 5000 nM by two-fold dilution, for 48 h.
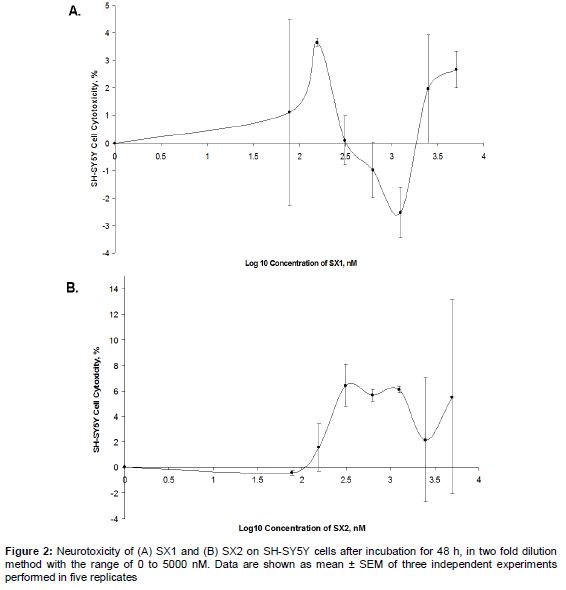
Cells were then rinsed twice with phosphate buffer saline (PBS) and 20 μL of 3-(4,5-dimethylthiazol-2-yl)-2.5-diphenyltetrazolium bromide (MTT) reagent (Bio Basic Canada Inc., Canada) stock solution (5 mg/mL) was then added to each well. The cells were then incubated at 37 ºC for 4 h, before 100 µL of DMSO was added to solubilize the purple formazan crystals. After an hour of incubation, the absorbance was measured at the wavelength of 570 nm using a microplate reader (M200 Tecan, Switzerland). To determine the MNTD or ½ MNTD of SX1 and SX2, a graph of percentage of toxicity against the log10 concentration of compounds was plotted. MNTD was determined at the first x-intercept where the toxicity is 0 %, while half MNTD (HMNTD) was half of the MNTD selected.
For the determination of neuroprotection against paraquat and 6-OHDA-induced cell death, SH-SY5Y cells were grown at 8,000 cells/well, cultured and starved as mentioned above. On the third day, the cells were incubated with SX1 and SX2 at their MNTDs and HMNTDs for 24 h. On the fourth day, the cells were treated with 200 µM paraquat (stock solution dissolved in filter sterilized deionized water) or 100 µM 6-OHDA (stock solution dissolved in filter sterilized deionized water and supplemented with 0.02 % of ascorbic acid and 0.85 % of NaCl) in complete growth medium for another 24 h.
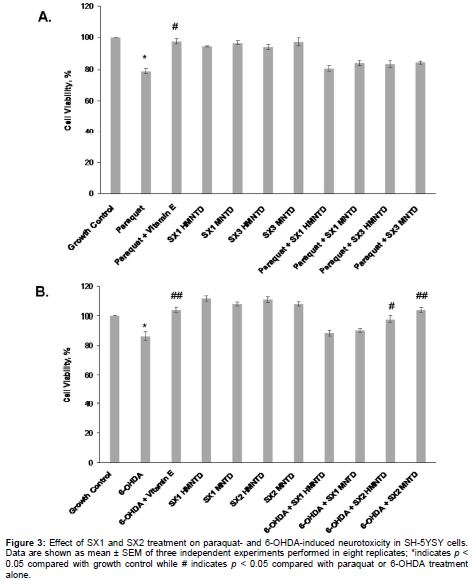
MTT assay was then performed as per described above and graphs of percentage cell viability vs. treatments were plotted. Growth control was represented by cells that were cultured with only the medium while the positive control of neuroprotection was represented by the addition of D-α-tocopherol (vitamin E; an antioxidant) during the treatment with paraquat or 6-OHDA.
Determination of intracellular free radical production by dichloro-dihydro-fluorescein diacetate (DCFH-DA) assay
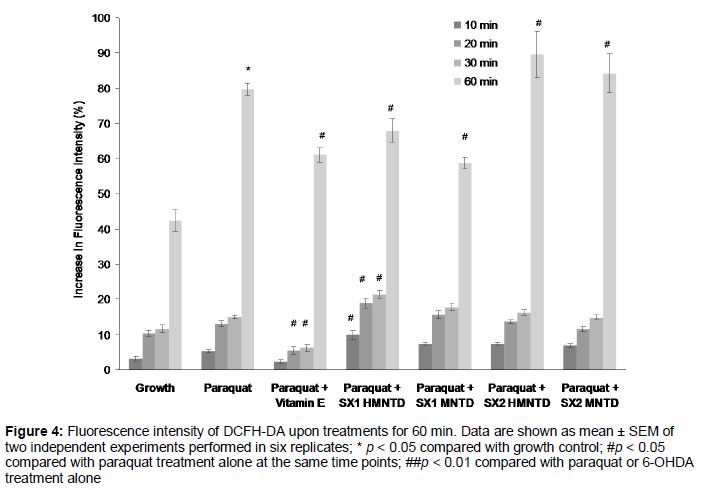
SH-SY5Y cells were grown at similar density, cultured and starved as mentioned above. On the third day, the medium was removed from each well and cells were rinsed with PBS twice. Then the cells were incubated with 100 µM of DCFH-DA (stock solution dissolved in filter sterilized deionized water) in complete growth medium at 37 °C in 5 % CO2 humidified incubator for 30 min. The growth medium containing DCFH-DA was then removed by pipetting, and the cells were rinsed twice with PBS and incubated with 200 µM paraquat or 100 µM 6-OHDA together with the presence or absence of SX1 and SX2 MNTD. Immediately, fluorescence was measured at 485 nm excitation wavelength and 530 nm emission wavelength using a microplate reader (M200, Tecan, Switzerland). The readings were taken every 10 min for 30 min and last reading was taken at 60 min of incubation time. The fluorescence readings were expressed in percentage change in fluorescence intensity compared with wells with media only and without cells.
Statistical analysis
Data was expressed as mean ± standard error of the mean (SEM) of at least two independent experiments, which were performed in replicates as stated in the figure legends. Statistical analysis was performed using Student’s t-test for comparison between means of two continuous variables using IBM SPSS Statistics software version 16.0 (IBM, NY, USA). A p-value of less than 0.05 was considered as statistically significant.
Results
Maximum non-toxic dose (MNTD) and half maximum non-toxic dose (½ MNTD)
Across the range of dosages of SX1 and SX2 tested on SH-SY5Y cells, neurotoxicity ranged from -3 to 7 % (); positive values indicate cell death, negative values indicate cell proliferation. Cells treated with SX1 showed more fluctuations in neurotoxicity across the range of dosages tested, while those treated with SX2 showed fluctuations above 0 %, starting from 100 nM. As determined from the graph, MNTDs of SX1 and SX2 were 1850 and 105 nM, respectively, while the HMNTDs were 925 and 52.5 nM, respectively. Overall, the MNTD and HMNTD of SX1 were observed to be 18-fold higher than SX2.
Neuroprotection against paraquat- and 6-OHDA-induced cell death
Treatment with paraquat alone significantly induced 21.03 ± 2.82 % of neurotoxicity on SH-SY5Y cells compared with growth control (A), while treatment with 6-OHDA alone induced 13.61 ± 2.96 % of neurotoxicity (B). Co-incubation of paraquat with vitamin E (positive control for neuroprotection) significantly increased the viability of SH-SY5Y cells by nearly 20 % as compared with cells treated with paraquat alone (A). Similarly, co-treatment of 6-OHDA with vitamin E significantly increased the viability of SH-SY5Y cells by nearly 18 % as compared with cells treated with 6-OHDA alone (B). Treatment with MNTDs and HMNTDs of SX1 or SX2 alone did not significantly increase the cell viability compared with the growth control (A and B). Co-incubation of MNTDs and HMNTDs of SX1 or SX2 with paraquat did not significantly protect the cells from neurotoxicity, compared with growth control (A). However, co-treatment of MNTDs and HMNTDs of SX2 (but not SX1) with 6-OHDA, significantly increased the cell viability by 11.55 ± 1.82 and 17.26 ± 1.87 %, respectively, compared with treatment with 6-OHDA alone (B). This indicates that SX2, but not SX1, had significant neuroprotection against 6-OHDA neurotoxicity, but not paraquat neurotoxicity.
Intracellular reactive oxygen species (ROS) production by cells treated with paraquat or 6-OHDA
As shown in , paraquat treatment alone over 60 min significantly induced 79.73 ± 4.09 % increment in intracellular ROS production in SH-SY5Y cells compared with untreated growth control. Addition of vitamin E (a potent antioxidant) to cells treated with paraquat significantly attenuated the production of intracellular ROS at 20, 30 and 60 min time points compared with paraquat treatment alone. Intracellular ROS production in cells treated with paraquat and SX1 and SX2 MNTDs and HMNTDs gradually increased across the time points, with SX1 HMNTD showing significances across all time points, compared with paraquat treatment alone ().
Addition of SX1 HMNTD significantly increased ROS production at earlier time points (10, 20, 30 min), but significantly attenuated approximately 15 % of intracellular ROS production at 60 min time point. SX1 MNTD also significantly did so at 60 min. However, treatment with SX2 HMNTD and MNTD significantly increased ROS production at 60 min, instead of decreasing it. The co-treatment of 6-OHDA with SX1 and SX2 HMNTDs and MNTDs produced similar results with the treatment with 6-OHDA alone (data not shown), indicating that SX1 and SX2 did not have any effect in attenuating or increasing intracellular ROS production in SH-SY5Y cells treated with 6-OHDA.
Discussion
Previously, it was shown that six newly isolated prenylated xanthones from ethyl acetate soluble extract of Cudrania tricusidata root bark (cudratrixanthone C, cudratrixanthone G, cudratrixanthone H, cudratrixanthone I, cudratrixanthone O, 3-O-methylcudratrixanthone G), and six other previously identified prenylated xanthones (cudraxanthone A, 6-deoxyiso-jacareubin, nigrolineaxanthone F, neriifolone A, cudraxanthone L, gerontoxanthone C), protected against 6-OHDA-induced cell death in human neuroblastoma SH-SH-SY5Y cells [8]. These compounds gave the EC50 values of 0.7 – 16.6 μM, compared with positive control curcumin - 6.0 μM. In our study, we used a slightly different approach in determining the neuroprotective property of two selected prenylated xanthones - 1,3,6-trihydroxy-9H-xanthen-9-one (SX1) and 1,3,6-trihydroxy-2-(3-methylbut-2-enyl)-9H-xanthen-9-one (SX2). We determined their HMNTDs and MNTDs first before these dosages were used to test for potential neuroprotection against paraquat and 6-ODHA-induced neuronal cell death. Determination of MNTD is crucial prior to subsequent neuroprotection assay so that the compound would not pose any neurotoxicity during treatment. This similar approach of MNTD determination first has been used in numerous neuroprotection assays of natural products, such as orientin [9] and edible bird nest (EBN) extract [10].
We found that the neurotoxicity of SX2 was two-fold more than the neurotoxicity of SX1, while the MNTD of SX1 was nearly 17 times higher than MNTD of SX2. Orientin, a flavonoid-like phytochemical which showed neuroprotection against oxidative stress-induced SH-SY5Y cell death, had MNTD of 20 µM, which was much higher than the MNTDs of SX1 and SX2 [9]. This discrepancy of toxicity profile of SX1 and SX2 is also evident in our previous studies, whereby the inhibitory concentration (IC50) of SX1 on HeLa and MDA-MB-231 were > 60 µM and > 60 µM, respectively [6], while IC50 of SX2 on HeLa and MDA-MB-231 were 48 µM and > 60 µM, respectively [7]. These differences in neurotoxicity profile of SX1 and SX2 could be due to the different mode of action by distinct degree of prenylation of xanthone [11].
Paraquat at 200 µM induced nearly 20 % of SH-SY5Y neuronal cell death over 24 h of incubation, consistent with the results of a previous study [12]. Paraquat-induced cell death is possibly caused by the oxidative stress generated during incubation. Specifically, new redox reaction between paraquat radical and oxygen generate true reactive oxygen species, O2- and H2O2, which significantly cause cell death [12,13]. However, 6-OHDA treatment caused only around 20 % of neuronal cell death compared to ≥50 % at similar concentration, as shown in previous studies [14,15]. 6-OHDA is prone to oxidation. Therefore, a prolonged incubation period of 24 h might have caused the 6-OHDA to be oxidized and degraded, rendering it to be no longer a bioactive neurotoxin [16].
Vitamin E as a lipophilic antioxidant attenuated paraquat and 6-OHDA-induced cell death to nearly 20 % of improved cell viability in SH-SY5Y cells, suggesting that lipid peroxidation plays a role in 6-OHDA and paraquat toxicity [17]. The co-incubation of paraquat with SX1 and SX2 HMNTDs and MNTDs did not significantly protect the cells against paraquat-induced neurotoxicity. In contrast, SX2 HMNTD and MNTD improved cell viability of 6-OHDA challenged SH-SY5Y cells. The majority of xanthones isolated in a previous study have relatively higher dose in order to improve cell viability upon introduction of neurotoxins, while only minor group of xanthones with relatively complex prenylation showed more potent neuroprotection at lower dose [8]. This suggests that the SX1 and SX2 are not as potent as natural xanthones isolated from Cudrania tricuspidata as the MNTDs of SX1 and SX2 were too low to provide neuroprotection against paraquat-induced cell death.
Cells treated with paraquat alone had the strongest intracellular ROS production over 60 min of treatment. This shows that neurotoxic paraquat ion (PQ+) accumulated in dopaminergic neuron to reestablish a new redox reaction intracellularly, leading to generation of intracellular free radicals such as superoxide and dopamine reactive substances in the neuron [3]. The co-incubation of paraquat with vitamin E significantly reduced intracellular ROS production at time point 10, 20, 30 and 60 minutes as compared to incubation with paraquat alone. A previous study demonstrated in rodent model that vitamin E could strengthen the antioxidative defense system with an increased activity of antioxidative enzymes, including superoxide dismutase (SOD) and glutathione peroxidase (GSH-px) [18].
The co-incubation of paraquat with SX1 HMNTD and MNTD attenuated intracellular ROS production but did not rescue the SH-SY5Y cells from paraquat-induced cell death, while SX2 HMNTD and MNTD maintained increased intracellular ROS production. The attenuation of intracellular ROS would normally rescue the cell from oxidative stress-induced cell death [10]. However, this was not observed in this study. One of the possible reasons is that the direct free radical scavenging did not sufficiently protect early apoptotic cells against neurotoxin-induced cell death. This indicates that there could other mechanisms involved other than direct anti-oxidation in rescuing paraquat-induced SH-SY5Y cell death. This is evident in a previous study, where the effective free radical scavenging activity of EBN extract did not rescue SH-SY5Y cells from oxidative stress-induced cell death [10].
Conclusion
1,3,6-Trihydroxy-2-(3-methylbut-2-enyl)-9H-xanthen-9-one (SX2) at 105 nM (MNTD) and 52.5 nM (HMNTD) concentrations successfully confer neuroprotection by significantly protecting SH-SY5Y cells against 6-OHDA-induced cell death by improving 10 and 17 % cell viability. Although intracellular ROS production was significantly attenuated by 1,3,6-trihydroxy-9H-xanthen-9-one (SX1) at 1850 nM (MNTD) and 925 nM (HMNTD) concentrations, the radical scavenging activity of the compounds did not significantly protect SH-SY5Y cells against paraquat-induced cell death.
Declarations
Acknowledgement
References
Archives


News Updates